
پروتئین غشائی درونی یک یا مجموعهای از پروتئینها ست که به طور دائمی به غشاء سلولی متصل شده است.
دو نوع پروتئین در غشاء وجود دارد: پروتئینهای انتگرال یا سرتاسری که در تمام ضخامت غشاء نفوذ میکنند و پروتئینهای محیطی که فقط به یک سطح غشاء میچسبند و در ان نفوذ نمیکنند. تعداد زیادی از پروتئینهای انتگرال کانال ها(یا منافذ) ساختاری ایجاد میکنند که از طریق آنها مولکولهای اب و مواد محلول درآب به ویژه یونها میتوانند بین مایع خارج سلولی و داخل سلولی انتشار یابند. این کانالهای پروتئینی دارا خواص انتخابی نیز هستند که دیفوزیون ترجیهی برخی مواد به میزان بیشتر از مواد دیگر را امکانپذیر میسازند. تعدادی دیگر از پروتئینهای انتگرال به عنوان پروتئینهای حامل برای انتقال دادن موادی عمل میکنند که در غیر این صورت نمیتوانستند از لایه دو طبقه چربی نفوذ کنند. گاهی نیز این پروتئینهای حامل مواد را در جهتی خلاف جهت انتشار طبیعی انها انتقال میدهند که « انتقال فعال» نامیده میشوند. تعداد بازهم دیگری از پروتئینها انتگرال به صورت انزیمها عمل میکنند.
چنین پروتئینهایی را تنها میتوان به کمک زدایندهها، حلالهای غیرقطبی، و یا عوامل واسرشتنده از غشاء سلولی جدا کرد.
غِشاء سلولی یا غشای پلاسمایی به پوسته نازک یاختهها که از یک دولایه فسفولیپیدی تشکیل شده گفته میشود.
غشای سلولی همچنین به عنوان مرز میان اندامکهای درون یاختهای عمل میکند. در واقع غشاء سلول جداری ست که محافظت از سلول را برعهده دارد. سیتو پلاسم در داخل غشاء سلول است و محیط خارج سلولی در خارج از آن قرار دارد. غشاء سلول از یک لایه پروتئین و دو لایه چربی تشکیل شده است. رفت و آمد مواد به داخل و خارج سلول از طریق کانالهای غشاء سلول انجام میشود. از بین رفتن این غشاء آسیب پذیری سلول را سبب میشود.
غشای پلاسمایی شامل دولایه فسفولیپیدی همراه با کلسترول و پروتئینهای درون غشایی یا سطح غشایی میباشد.
به بیانی دیگر بخش اعظم اندامکهای سلول بوسیله غشاءهایی مفروش شدهاند که به طور عمده از لیپیدها و پروتئینها تشکیل شدهاند.این غشاءها شامل غشاء سلولی، غشاء هسته، غشای رتیکولوم اِندوپلاسمیک و غشاء میتوکندریها، لیزوزومها و دستگاه گلژی هستند.
لیپیدهای غشاءها سدی ایجاد میکنند که از حرکت آزاد آب موجود در مایع میانیاخته و مواد محلول در آب از یک بخش سلول به یک بخش دیگر جلوگیری میکنند زیرا آب در چربی محلول نیست. اما باید دانست که ملکولهای پروتئینی در غشاء٬ غالباً در سراسر عرض غشاء نفوذ کرده و به این ترتیب مسیرهای اختصاصی، که غالبا منافذ یا Pores نامیده میشوند برای عبور مواد ویژه از غشاء به وجود میآورند. همچنین بسیاری از سایر پروتئینهای غشاء آنزیمها هستند که تعداد زیادی از واکنشهای شیمیایی مختلف را کاتالیز میکنند.
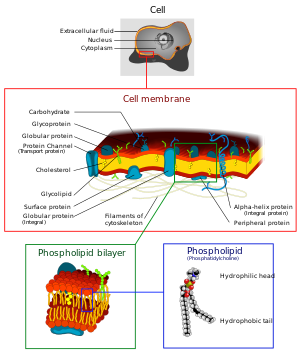
غشاء سلول
غشای سلول که به طور کامل سلول را احاطه میکند، یک ساختار خم پذیر ارتجاعی نازک به ضخامت ۵/۷ تا ۱۰ نانومتر است. غشاء تقریباً از پروتئینها و لیپیدها تشکیل شدهاست و ترکیب تقریبی عبارت است از: پروتئینها ۵۵ درصد، فسفولیپیدها ۲۵ درصد، کلسترول ۱۳ درصد و سایر لیپیدها ۴ درصد و کربوهیدراتها ۳ درصد.
سد لیپیدی غشای سلول از نفوذ آب جلوگیری میکند: ساختار پایه غشای سلول یک لایه چربی دو طبقهاست که یک ورقه نازک از لیپیدها فقط به ضخامت دو مولکول بوده و در سراسر سطح سلول٬ یکپارچهاست. جایجای این ورقه نازک لیپیدی، مولکولهای پروتئینی درشت از نوع کروی شکل قرار دارند.
ساختارپایه لایه دوطبقه چربی از مولکولهای فسفولیپید تشکیل شدهاست. یک انتهای هر مولکول فسفولیپید در آب محلول بوده یعنی آبدوست (هیدروفیل) است. انتهای دیگر فقط در چربیها محلول بوده یعنی آبگریز (هیدروفوب) است. انتهای فسفاتی فسفولیپد هیدروفیلیک و اسید چربی آن هیدروفوبیک است. چون قسمتهای آب گریز فسفولیپیدها به وسیله آب دفع میشوند اما به سوی یکدیگر جذب میشوند لذا دارای یک تمایل طبیعی هستند و آنها در پهلوی همدیگر در مرکز غشاء قرار دارند. بخشها فسفاتی آب دوست دو سطح غشاء را که در تماس با اطراف است میپوشانند. لایه دو طبقهٔ چربی در وسط غشاء به مواد طبیعی محلول در آب از قبیل یونها، گلوکز و اوره نفوذ پذیر است بر عکس، مواد محلول در چربی از جمله اکسیژن، کربن دی اکسید و الکل میتوانند با سهولت در این بخش از غشاء نفوذ کنند. یک صفت ویژه لایه دوطبقه چربی ان است که یک مایع است نه یک جامد. بنابر این بخشهایی از غشاء میتوانند عملاً در سطح غشاء از یک نقطه به یک نقطه دیگرجریان پیدا کنند. پروتئینها و سایر مواد محلول در غشای دو طبقه لیپیدی یا شناور در آن تمایل دارند که به کلیه غشای سلول انتشار یابند.
مولکولهای کلسترول درغشا نیز ماهیت چربی دارند زیرا هسته استروئیدی آنها بسیار محلول در چربی است. این مولکولها از یک نظر در لایه دو طبقه غشاء حل شدهاند. این مولکوها به طور عمده به تعیین مزان نفوذ پذیری لایه لایه دو طبقه به اجزای محلول در آب و مایعات بدن کمک میکنند. کلسترول همچنین قسمت زیادی از قابلیت تحرک غشا را کنترل میکند.
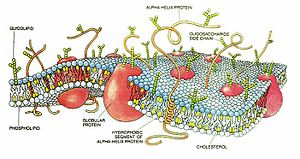
پروتئینهای غشای سلول
تودههای کروی شکل که در لایه دو طبقه چربی شناورند، اینها پروتئینهای غشاء هستند که قسمت اعظم آنهارا گلیکو پروتئینها تشکل میدهند. دو نوع پروتئین در غشاء وجود دارد: پروتئینهای انتگرال یا سرتاسری که در تمام ضخامت غشاء نفوذ میکنند و پروتئینهای محیطی که فقط به یک سطح غشاء میچسبند و در آن نفوذ نمیکنند.
تعداد زیادی از پروتئینهای انتگرال کانال ها(یا منافذ) ساختاری ایجاد میکنند که از طریق آنها مولکولهای آب و مواد محلول درآب به ویژه یونها میتوانند بین مایع خارج سلولی و داخل سلولی انتشار یابند. این کانالهای پروتئینی دارای خواص انتخابی نیز هستند که دیفوزیون ترجیحی برخی مواد به میزان بیشتر از مواد دیگر را امکانپذیر میسازند. تعدادی دیگر از پروتئینهای انتگرال به عنوان پروتئینهای حامل برای انتقال دادن موادی عمل میکنند که در غیر این صورت نمیتوانستند از لایه دو طبقه چربی نفوذ کنند. گاهی نیز این پروتئینهای حامل مواد را در جهتی خلاف جهت انتشار طبیعی انها انتقال میدهند که « انتقال فعال» نامیده میشوند. تعداد بازهم دیگری از پروتئینها انتگرال به صورت آنزیمها عمل میکنند.
پروتئینهای محیطی به طور عمده روی سطح داخلی غشاء وجود دارند و غالبا به یکی از پروتئینهای انتگرال چسبیدهاند. این پروتئینهای محیطی تقریبا به طور کامل به صورت آنزیمها یا سایر انواع کنترل کنندهها عمل داخلی سلولی عمل میکنند.

کربوهیدراتهای غشاء – گلیکوکالیس سلولی [ویرایش]
کربوهیدراتهای غشاء تقریبا همیشه به صورت ترکیب با پروتئینها به شکل گلیکوپروتئینها و گلیکولیپیدها وجود دارند. در واقع بخش بزرگی از پروتئینهای انتگرال از نوع گلیکوپروتئینها و در حدود یکدهم مولکولهای لیپید از نوع گلیکولیپیدها هستند. بخشهای گلیکو در این مولکولها تقریبا همیشه به طرف سطح خارجی سلول برآمدگی پیدا میکنند و از سطح سلول به سوی خارج آویزان هستند. بسیاری از ترکیبهای کربوهیدراتی دیگر موسوم به پروتئوگلیکانها که به طور عمده از مواد کربوهیدراتی تشکیل شدهاند که به هستههای کوچک پروتئینی متصل شدهاند نیز غالباً به طور سست به سطح خارجی سلول متصل هستند. بهاین ترتیب تمامی سطح سلول دارای یک پوشش سست کربوهیدراتی موسوم به گلیکوکالیس است.
این بخشهای کربوهیدراتی که به سطح خارجی سلول متصل شدهاند دارای چندین عمل مهم هستند:
1.بسیاری ازانها بار الکتریکی منفی دارند و این موضوع به بیشتر سلولها یک سطح با بار کلی منفی میبخشد که سایر اشیای منفی را دفع میکند.
2.گلیکو کالیس بعضی از سلولها به گلکو کالیس سلولهای دیگر میچسبند و به این ترتیب سلولها را به یک دیگر میچسبانند.
3.بسیاری از کربوهیدراتها به عنوان مواد حامل برای گرفتن هورمونهایی ازجمله انسولین عمل میکنند. و پس از انجام این عمل این مجموعه پروتئینهای چسبیده به سطح داخلی غشاء را فعال میکند که به نوبه خود یک سری متوالی از انزیمهای داخل سلولی را فعال میکنند.
4.بعضی از بخشهای کربوهیدراتی وارد واکنشهای شیمیایی میشوند.
محیط درونی سلول از بیرون آن متفاوت است. این اختلاف در تمام مدت زیست سلول بوسیله یک غشای نازک که سطح سلول را میپوشاند که آن را غشای سلولی یا غشای سیتوپلاسمی میگویند که ورود و خروج مولکولها و یونها از کنترل میکند.
نگاه اجمالی
غشای سلولی ساختمانی است به ضخامت 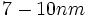 که محدوده سلول را معین کرده و به عنوان سد انتخابی ، مبادله مواد بین سلول و محیط اطرافش را کنترل میکند. غشا از دو لایه تقریبا ممتد لیپیدی ساخته شده که در آنها مجموعههای پروتئینی بطور پراکنده وارد شدهاند علاوه بر این پروتئینهای غشایی پروتئینهای دیگری که از نوع پروتئینهای حاشیهای هستند، در غشای دو لایه و اغلب روی سطح داخلی قرار میگیرد. بنابراین غشا بسیار نامتقارن است. بخشی از عدم تقارن غشا مربوط به زنجیرههای الیگوساکاریدی میباشد که تنها به سطح خارجی غشا چسبیدهاند.
که محدوده سلول را معین کرده و به عنوان سد انتخابی ، مبادله مواد بین سلول و محیط اطرافش را کنترل میکند. غشا از دو لایه تقریبا ممتد لیپیدی ساخته شده که در آنها مجموعههای پروتئینی بطور پراکنده وارد شدهاند علاوه بر این پروتئینهای غشایی پروتئینهای دیگری که از نوع پروتئینهای حاشیهای هستند، در غشای دو لایه و اغلب روی سطح داخلی قرار میگیرد. بنابراین غشا بسیار نامتقارن است. بخشی از عدم تقارن غشا مربوط به زنجیرههای الیگوساکاریدی میباشد که تنها به سطح خارجی غشا چسبیدهاند.
لیپیدهای غشا
لیپیدهای غشایی شامل فسفولیپید (فسفوگلیسرید و اسفنگولیپید) و کلسترول میباشد. فسفولیپیدها مولکولهایی هستند که از یک قسمت سر مانند و یک دنباله متصل به آن تشکیل شدهاند. قسمت سری که به سر قطبی Polar head نیز موسوم است، حاوی گروه فسفات بوده و آب دوست Hydropgilic میباشد قسمت دنباله از دو زنجیره اسید چرب تشکیل شده و آب گریز Hydrophobic میباشد. دنباله غیر قطبی Non polartail نیز نامیده میشود.
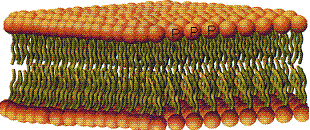
فسفولیپیدها در این ساختمان دولایه به ترتیبی است که قطبهای هیدروفیل آنها در سطح داخلی و خارج سیتوپلاسم و دنبالههای هیدروفوب آنها در مرکز قرار گرفته است و همین امر باعث سه لایه دیده شدن غشا با میکروسکوب الکترونی میگردد. از دیگر لیپیدهای غشایی ، کلسترول میباشد که در حد فاصل اسیدهای چرب قرار گرفته است. میزان سیالیت غشا بستگی به میزان کلسترول آن دارد. هرچه کلسترول بیشتر سیالیت غشا نیز بیشتر خواهد بود.
پروتئینهای غشا
پروتئینها که در اکثر غشاها بیش از 50 درصد وزن آن را تشکیل میدهند، دارای وظایف ساختمانی مانند حفظ شکل سلول مانند گویچههای قرمز خون و عملکری (مثل فعالیت آنزیمی) متعدد میباشند. این پروتئینها به دو صورت محیطی percpheral و سراسری یا داخلی Integral protein دیده میشوند و انواع آنها در ارگانلها و سلولهای مختلف میتواند متفاوت باشد.
انواع پروتئینهای غشا
•پروتئینهای محیطی : در سطح غشا قرار دارند و بسیاری از آنها دارای فعالیت آنزیمی میباشند.
•پروتئینهای انتگرال : پروتئینهای درشت مولکولی هستند که مستقیما در داخل لیپید دو لایه قرار گرفتهاند. اندازه این پروتئینها به حدی است که سراسر ضخامت لیپید دولایه را طی میکنند و در هر دو سطح غشا نمایان هستند و یا اینکه تا حدی در ضخامت لیپید دو لایه فرو رفتهاند و فقط در سطح داخلی یا خارجی غشا نمایان میباشند. از آنجا که مواد محلول در آب قادر به عبور از لیپید دولایه نمیباشند عقیده بر این است که پروتئینهای سراسری به عنوان کانالهایی برای مبادله مواد محلول در آب از قبیل یونها عمل میکنند.
کربوهیدراتهای غشا
کربوهیدراتهای غشا از نوع الیگوساکاریدها میباشند. الیگوساکاریدها به کربوهیدراتهای متشکل از چند واحد قندی اطلاق میگردد. الیگوساکاریدها عمدتا در سطح خارجی غشا و متصل با پروتئینها و لیپیدها یعنی به صورت گلیکوپروتئین و گلیکولیپید دیده میشوند. ترکیبات فوق هم دارای خاصیت آنتی ژنیک میباشند و هم به عنوان رسپتور (گیرنده) در سطح سلول عمل میکنند. وجود رسپتور در سطح سلول باعث میشود که مواد معینی بتوانند وارد سلول شوند و یا سلول نسبت به هورمون معینی که رسپتور آن را دارد عکسالعمل نشان دهد.
سیستمهای انتقال از غشا
انتشار
مبادله مواد محلول در چربی ، آب ، گاز اکسیژن و دیاکسید کربن بین سلول و محیط اطراف انتشار نامیده میشود. در صورتی که انتشار مواد با اتصال به مولکولهای دیگر تسریع گردد آن را انتشار تسهیل شده مینامند. چون انتشار تسهیل شده با دخالت پروتئینهای انتگرال صورت میگیرد. پروتئینهای دخیل در این امر را حامل Porter یا انتقال دهنده گویند.
انتقال فعال Active transport
نقل و انتقال الکترولیتها ( ) بین سلول و محیط اطراف آن اگر بر خلاف شیب غلظت و با صرف انرژی انجام میگیرد.
) بین سلول و محیط اطراف آن اگر بر خلاف شیب غلظت و با صرف انرژی انجام میگیرد.
آندوسیتوز Endocytosis
•پینوسیتوز : در این روش که به آشامیدن سلول نیز موسوم است ابتدا مایعات و مواد محلول و بسیار ریز به رسپتورهای غیر اختصاصی سطح سلول متصل میشوند سپس غشا در آن ناحیه فرو رفته شده و به تدریج با عمق رشد ، فرورفتگی و بهم چسبیدن لبههای آن قسمت فرو رفته به صورت وزیکول در آمده و از غشای سلول جدا شده و در سیتوپلاسم رها میگردد. این وزیکول ممکن است به لیزوزوم پیوسته و تحت تاثیر آنزیمهای آن قرار گیرد و یا به عنوان حامل عمل کرده و پس از طی بخش داخلی سلول و پیوستن به غشای مقابل محتویات خود را از سلول عبور میدهند. عبور مواد از دیواره مویرگها نمونهای از این روش میباشد.
•آندوسیتوز با واسطه رسپتور : این روش انحصارا برای ورود موادی معین درون سلولهایی معین مورد استفاده قرار میگیرد، نیازمند اتصال ماده با رسپتور اختصاصی مربوطهاش در سطح سلول میباشد. برخی از هورمونها و برخی ویروسها به این طریق وارد سلول میشوند.
•فاگوسیتوز: فاگوسیتوز در مقایسه با آندوسیتوز با واسطه رسپتور ، روشی غیر اختصاصی است.
سلولهای معینی مانند ماکروفاژها با استفاده از این روش ، باکتریها و قارچهای وارد شده به بدن و یا حتی سلولهای آسیب دیده و فرسوده را فاگوسیتوز میکنند.
اگزوسیتوز
برعکس آندرسیتوز در عمل اگزوسیتوز مواد از محیط داخل سلول به خارج از سلول انتقال مییابند. این مواد که شامل ذرات ترشحی ساخته شده در سلول و یا مواد باقیمانده حاصل از تجزیه لیزوزوم میباشند به صورت وزیکول ترشحی یا دفعی دیده میشوند. پس از چسبیدن وزیکول ترشحی یا دفعی به غشای سلول ، غشا در محل چسبیدگی از بین میرود و به این طریق محتویات وزیکول به خارج از سلول تخلیه میگردد.
وظایف غشای سلولی
1.حفظ شکل مشخص سلول و جلوگیری از خروج محتویات آن. این عمل برای پردهای که فقط 75 آنگستروم ضخامت دارد بسیار عجیب و ناباورانه است. اگر غشای سلولی در محلی پاره شود، سیتوپلاسم از آن محل خارج میشود و سلول میمیرد.
2.جلوگیری از خروج مواد لازم برای سلول و وارد کردن موادی که سلول لازم دارد. این غشا مانند یک نگهبان جلوی عبور مواد ممنوع الخروج یا ممنوع الورود را میگیرد و تنها آنهایی را که لازم است، وارد سلول میکند. موادی که وارد سلول میشوند دو گروه هستند: یک گروه بطور عادی وارد سلول میشوند، بعنی از آنها که مقدار آنها در خارج سلول بیشتر است، به داخل آن منتشر میشوند. گروه دیگر نحوه ورودشان بسیار جالب است.
زیرا ممکن است مقدار آنها در داخل سلول چندین برابر بیرون باشد و ظاهراً باید از آن خارج شوند، ولی در جدار غشای سلولی موادی وجود دارد که آنها را به داخل میبرد. این مواد شیمیایی ، مانند مورچههایی که دانههای گندم و سایر مواد غذایی را میگیرند و به داخل لانه خود میبرند، به موادی که باید به داخل سلول برده شود میچسبند و سپس همراه آنها از غشای سلولی عبور میکنند، ولی قبل از رسیدن به سیتوپلاسم ، ماده مزبور را رها کرده و آن را با فشار وارد سیتوپلاسم میکنند و خود فوراٌ برای آورن طعمه جدید به طرف خارج غشا میروند. مواد شیمیایی دیگری نیز وجود دارند که همین عمل را در مورد خارج کردن موادی که سلول لازم ندارند، انجام میدهند.
1) واکنش های آگلوتیناسیون :
به هم چسبیدن میکروارگانیسم ها و بعضی از کلاس های آنتی بادی ایجاد شده بر علیه آنها. به این آنتی بادی ها "آگلوتینین" و به این خاصیت "آگلوتیناسیون" گفته می شود.
2)واکنش های پرسی پیتاسیون:
رسوب بعضی از آنتی بادی ها و ملکول های مواد محلول ( آنتی ژن محلول).به آنتی بادی "پرسیپیتین" و این واکنش را "پرسی پیتاسیون " میگویند.
3) واکنش های فلوکولاسیون:
وقتی آنتی ژن به صورت ذرات کلوئیدی باشد (مثل کاردیولیپین قلب در تست VDRL ) واکنش را "فلوکولاسیون" نامند.
4) واکنش های هماگلوتیناسیون:
اگر آنتی ژن غیرمحلول گلبول قرمز باشد واکنش بین گلبول های قرمز و آنتی بادی ضد آنرا "هماگلوتیناسیون" می نامند.
5) Themass action theory :
اتصال آنتی ژن به آنتی بادی اختصاصی و دوطرفه است و از قوانین تئوری عکس العمل بین اسیدهای ضعیف و بازهای ضعیف پیروی می کند.
6) غلظت اپتیمم آنتی بادی و آنتی ژن :
غلظتی از آنتی ژن و آنتی بادی که موجب حداکثر واکنش سرولوژی می گردد.
7) پدیده prozone :
بیشتر بودن مقدار آنتی بادی نسبت به آنتی ژن
8) پدیده post zone:
کاهش مقدار آنتی بادی متصل به آنتی ژن
9) آنتی بادی هتروفیل:
نوعی آنتی بادی اکثرا از کلاس IgM که با آنتی ژن های متنوعی از منابع مختلف واکنش میدهد.( در بیماری منونوکلئوز عفونی)
10) آگلوتینین های سرد:
نوعی اتو آنتی بادی از جنس گلیکو پروتئین و یا گلیکو لیپید از کلاس IgM که با بعضی از آنتیژن های گروه خونی در سطح گلبول های قرمز در حرارت های پایین تر از دمای بدن واکنش می دهند.(در سیستم آنتی ژنی I/i)
11) کرایو گلوبولین ها :
پروتئین های سرمی اکثرا از نوع ایمونوگلوبولین ها کی بصورت قابل برگشت در درجات پایین رسوب می کند.
12) آزمایش کومبس:
از این تست برای شناسایی آنتی بادی ها با قدرت اتصال به آنتی ژن ولی فاقد قدرت آگلوتیناسیون استفاده می شود. این آنتی بادی ها ناقص یا مصدود کننده می باشند.
13) فیکساسیون کمپلمان:
مصرف کمپلمان جهت تعیین و اندازه گیری آنتی بادی ها- آنتی ژن ها یا هردو
14) آرتریت روماتوئید:
نوعی بیماری مزمن کمپلکس ایمنی موضعی و جزو بیماری های خودایمنی میباشد که با تست RF مثبت همراه است.
15) پروتئین های فاز حاد:
بر اثر ضایعات بافتی- نکروز- التهاب- عفونت ها- اعمال جراحی یا سرطان ها در سرم و پلاسما ایجاد می شود. مثل CRP
16) تست ASO (آنتی استرپتولیزین O):
تعیین تیتر آنتی بادی ایجاد شده علیه استرپتولیزین O از باکتری استرپتوکک. اساس تست خنثی سازی آنزیم میباشد.تیتر بالا تر از Todd 200 نشانه ی بیماری است.
17) بیماری منونوکلئوز عفونی:
نوعی بیماری ویروسی توسط ویروس اپشتن بار (EBV) که تست تشخیصی آنPaull-Bunnell میباشد.
18) Davidsoin Test:
آزمایش تاییدی پس از مثبت شدن آزمایش پال-نوبل
19) آزمایش ویدال:
برای تشخیص بیماری حصبه (تیفوئید) و شبه حصبه (پاراتیفوئید) استفاده میشود. در 90% تا 95% مبتلایان به سالمونلا از هفته چهارم به بعد تست مثبت میشود. وجود تیتر آنتی بادی 80/1در برابر آنتی ژن O وتیتر 40/1در برابر آنتی ژن H فرد مشکوک به بیماری می باشد. تیتر Vi در حاملین سالم بیشتر از دو آنتی ژن دیگر است.
20) آزمایش رایت:
تست سرولوژیک برای شناسایی بروسلوز علاوه بر این تست از تست 2MEنیز در تشخیص این بیماری استفاده می شود.
21) آزمایش وایل- فلیکس:
سزم مبتلایان به بیماری تب تیفوسی سوش هایی از باکتری پروتئوس را به شدت آگلوتینه میکند.
22) راژین سفلیس:
نوعی آنتی بادی که در مبتلایان به سفلیس ایجاد می شود تست های تشخیصی آن (RPR (Rapid Plasma Reagin و (VDRL (Veneral Disease Research Laboratoryو( ART (Automated Reagin Test میباشد.
23) آنتی بادی دونات لنداشتاینر:
نوعی آنتی بادی در مرحله سوم بیماری سفلیس خصوصا فرم مادرزادی ایجاد میشود و از گروه آگلوتین های سرد می باشد.
24) تست کولمر یا ثبوت مکمل رایتر:
تستی برای شناسایی آنتی بادی گروه ترپونما بوده ولی تست اختصاصی این بیماری نیست. جواب منفی آن دارای ارزش زیادی میباشد.
25) تست بی حرکت کردن ترپونما پالیدوم:
تست اختصاصی شناسایی سفلیس بوده . سوش مصرفی دراین تست سوش نیکلس میباشد.
26) تست پوستی توبرکلین:
در این تست از PPD یا پروتئین خالص شده مایکو باکتریوم ها استفاده می شود . رایج ترین تست پوستی توبرکولین تست مانتو می باشد .
27) تست پوستی لپرومن:
جهت بررسی ایمنی افراد نسبت به مایکو باکتریوم لپره
28) تست فوشای:
تست پوستی تاخیری کاملا اختصاصی در تشخیص تولارمی
29) تست لیشمانین یا مونته نگرو:
تست پوستی که در مطالعات اپیدمیولوژیکی و انجام واکسیناسیون و تشخیص بیماری به کار میرود
30) Kveim test:
تست پوستی تاخیری جهت شناسایی بیماران مبتلا به سارکوئیدوز
31) تست پوستی کازونی:
تست پوستی فوری در تشخیص بیماری کیست هیداتید (نوعی بیماری با انگل اکینو کوکوس)
32) تست پوستی شیک:
تعیین مصونیت افراد در برابر بیماری دیفتری
3۳) تست پوستی دیک:
جهت بررسی مقاومت افراد نسبت به بیماری مخملک
3۴) تست پوستی شولتز- شارلتون:
تشخیص مخملک
ریشه لغوی
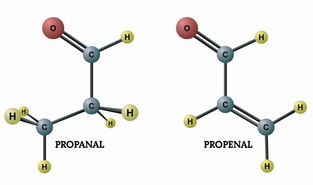
کلمه آلدئید (Aldehyde) از دو واژه الکل(Alcohal) و هیدروژنگیری (dehydrogenation) گرفته شده است. هرگاه از الکل نوع اول هیدروژنگیری شود، در آن صورت ، ماده ای تولید میگردد که آلدئید نامیده میشود.
نامگذاری آلدئیدها
در نامگذاری قدیمی ، نام آلدئید از نام اسید آلی مربوطه آن گرفته میشد. مثلا آلدئید فرمیک از اسید فرمیک ، آلدئید استیک را از اسید استیک و آلدئید پروپیونیک از اسید پروپیونیک بوجود آمده است. در نامگذاری جدیدی برای نامگذاری آلدئیدها ، ابتدا طولانیترین زنجیر حاوی عامل آلدئیدی را انتخاب میشود و سپس شماره گذاری اتمهای کربن از جهتی که گروه آلدئیدی قرار گرفته است، انجام میگیرد. برای نامگذاری ، ابتدا شماره و نام شاخههای فرعی را نوشته ، در پایان ، اسم هیدروکربن را بطور کامل ذکر و پسوند آل (al) بر آخر آن افزوده میشود.
تهیه آلدئیدها از اکسید گردن الکلهای نوع اول
با استفاده از برخی اکسید کنندههای ملایم مثل دیاکسیدمنگنز ، واکنشگر CrO3/H+/CH3COCH3) Jones) یا واکنشگر CrO3/HCl/Pyridine) Sarett) یا واکنشگر (CrO3/2pyridine/CH2Cl2) و کرومیل کلرید Cr2Cl2 ، الکل های نوع اول و دوم بصورت محدود اکسید میشوند.
الکل های نوع اول به آلدئید و نوع دوم به کتون هم کربن خود تبدیل میگردند.
تهیه آلدئیدها و کتونها با استفاده از ترکیبات آلی فلزی
گاهی اوقات از ترکیبات آلی فلزی برای سنتز آلدئیدها و کتونها استفاده میشود. در سالهای اخیر ، برای سنتز ترکیبات کربونیلدار از R2Cd یا RZnX یا R2Zn استفاده زیاد شده است. قدرت هسته خواهی ملایم و محدود این واکنشگرها این امکان را فراهم میکند که از کلرواسیدها ، کتونهای مربوطه سنتز شود.
تهیه آلدئیدها از اورگانومنیزینها
از فعل و انفعال اورگانومنیزینها با نیتریلها و آمیدها نیز میتوان آلدئید و کتون تهیه نمود. البته برای تهیه آلدئید باید آمید به شکل فرم آمید باشد.
سنتز آلدئیدها از واکنش آلکوکسی هیدریدلیتیم آلومینیوم با آمیدها و کلرواسیدها
احیای کلرواسیدها و آمیدها به کمک هیدریدیهای مناسب از قبیل تریآلکوکسی هیدرید لیتیم آلومینیوم به تشکیل آلدئید مربوطه منجر میگردد. توجه به این نکته ضرورت دارد که احیای کلرواسیدها بوسیله تریترسیو بوتوکسی لیتیم آلومینیوم در دمای پایین در حدود 78- درجه سانتیگراد و در حلالهای مناسب مانند دیگلایمMeO-CH2-CH2-O-CH2-CH2-OMe انجام میشود.
برای تبدیل آمیدها به آلدئیدهای مربوط ، از دیآلکوکسی هیدرید لیتیم آلومینیوم نیز میتوان استفاده کرد.
تهیه آلدئیدها از طریق هیدروفرمیه کردن آلکنها
آلکنها در حضور کاتالیزور تریستری فنیل فسفین رودیم کلراید با هیدروژن و منوکسید کربن ترکیب و به تشکیل آلدئید منجر میشود. با توجه به اینکه در این واکنش به یکی از کربنهای پیوند دوگانه ، هیدروژن و به کربن دیگر ، عامل آلدئیدی (-CHO) متصل میشود، لذا این واکنش را هیدروفرمیله شدن میگویند. در حضور کاتالیزور ذکر شده فقط یک نوع آلدئید تولید میشود. هر گاه از اکتا کربونیل دیکبالت به عنوان کاتالیزور استفاده شود، دو نوع ایزومر تولید میشود.
تهیه آلدئیدها از نیتریلها
در دمای پایین ، نیتریلها با هیدریدلیتیم آلومینیوم ترکیب میشوند و کمپلکس حد واسط را تشکیل میدهند. این کمپلکس در حضور محلول اسید هیدرولیز میشود و آلدئید مربوطه را تولید میکند.
سنتز آلدئیدهای آروماتیک با استفاده از واکنش کاترمن
یکی از روشهای تهیه آلدئیدهای آروماتیک این است که در حضور یک اسید لوئیس مناسب مثل کلروآلومینیوم یا کلرید روی ، سیانید هیدروژن و کلرید هیدروژن با ترکیبات آروماتیک واکنش داده شوند.
سنتز آلدئیدهای آروماتیک با استفاده از دیمتیل فرمآمید فسفریل کلراید
از اختلاط اکسی تریکلرو فسفر با فرمآمید ، کمپلکس تشکیل میشود که به دیمتیل فرمآمید فسفریل کلراید موسوم است و میتواند در واکنشهای جانشینی الکترونخواهی آروماتیک شرکت نماید. با این روش حتما آلدئیدهای هتروسیکلهایی مانند تیوفن و فوران نیز سنتز شدهاند.
سنتز آلدئیدهای آروماتیک از فنل ها و کلروفرم در حضور هیدروکسید سدیم
با استفاده از واکنش کلروفرم قلیایی شده با فنلها و از طریق جانشینی الکترونخواهی دیکلرو کاربن روی حلقه آروماتیک میتوان آلدئید سنتز نمود.

استالدئید
خواص شیمیایی آلدئیدها و کتون ها
آلدئیدها و کتونها در چند نوع فعل و انفعال شرکت میکنند که اهم آنها به قرار زیر است:
•حمله الکترونخواهی اسیدهای لوئیس روی اکسیژن گروه کربونیل ، موجب افزایش دانسیته بار مثبت کربن گروه کربونیل میشود که در نهایت ، موجب افزایش خصلت اسیدی پروتونهای کربنهای آلفای کربونیل میگردد. صحت این نکته بوسیله روشهای افزاری تایید شده است. به عنوان مثال ، محلولی از استن و سوپراسید (SbCl5 , FSO3H ) در دیاکسید سولفور مایع و در60- درجه سانتیگراد بوسیله n.m.r مطالعه و مشاهده شده است که جذب پروتونهای استن در میدانهای ضعیفتری صورت میگیرد.
•حمله هستهخواهی بر کربن گروه کربونیل ، دومین دسته وسیع از واکنشهای آلدئیدها و کتونها را تشکیل میدهد. به عنوان مثال ، از افزایش آب بر آلدئیدها و کتونها ، دیول دو قلو (gemdiol) ایجاد میشود و درصد تشکیل آن به ساختمان ماده و به پایداری محصول حاصل بستگی دارد. به عنوان مثال ، مقدار دیول دو قلوی حاصل از استون در دمای 20 درجه سانتیگراد خیلی کم و قابل اغماض میباشد، درصورتی که آلدئید فرمیک و تریکلرو استالدئید بخوبی و بطور کامل به دیول دوقلو مربوط به تبدیل میشوند.
ترکیبهای اسیدهای کربوکسیلی به فرمول R--COOR، استر نامیده میشود که در آن گروه -R یک گروه آلکیل یا آریل میباشد و یک گروه عمده از ترکیبات آلی را تشکیل میدهند. استرهای حلقهای نیز که لاکتون نامیده میشوند، جزو گروه استرها میباشند.
یک نوع واکنش برای تولید استر

خواص استر
استرها غالبا فرار و معطرند و برخی از آنها در میوههای رسیده یافت میشوند. مثلا استات ایزوپنتیل ، بوی موز است، والرات ایزوپنتیل بوی سیب بوده و پروپیونات ایزوبوتیل ، بوی نیشکر است.
موارد استفاده از استر
خیلی از استرها مانند استات اتیل و استات بوتیل ، بعنوان واکنشگر و یا حلال و نرم کننده رزینها در آزمایشگاهها و صنعت مورد استفاده قرار میگیرند.
استرهای طبیعی
مومها
چربیها و روغنها و مومهایی که در طبیعت یافت میشوند، حاوی استرهایی با جرم مولکولی بالا میباشند که به لیپید موسومند. مومها مخلوط پیچیدهای از استرها ، الکلها و آلکانهای با زنجیر طویل میباشند، ولی جزء اصلی تشکیل دهنده آنها ، استرهایی میباشند که از واکنش اسیدهای چرب و الکلهای با زنجیر طویل بوجود میآیند.
از جداسازی و مطالعه مواد تشکیل دهنده موم زنبور عسل معلوم شده است که میریسیل پالمیتات ، بیشترین مقدار آن را تشکیل میدهد که یک استر است.
چربیهای جامد و روغنهای مایع
چربیهای جامد و روغنهای مایع ، استرهایی هستند که از واکنش اسید چرب سنگین و گلسیرین بوجود میآیند و گلیسیرید نامیده میشوند. تعداد کربن اسیدها بین هشت تا بیست و دو میباشد. مطالعات نشان داده است که اسیدهای موجود در استرها ممکن است از یک نوع نباشند و بطور اتفاقی روی گلیسیرین قرار گرفته باشند. بعنوان مثال ، یک مولکول گلسیرین ممکن است به سه گروه استئارات یا یک مولکولپالمتیات و دو مولکول استئارات و … متصل شده باشد.
مهمترین اسیدهای چرب اشباع شدهای که از هیدرولیز چربیها و روغنها بدست آمدهاند، عبارتند از: اسید لوریک (Lauric acid) ، اسید پالمتیک (Palmitic acid) ، اسید استئاریک (Stearic acid). روغنهای مایع به مقدار زیاد ، گلیسیریدهای اسیدهای چرب اشباع نشده هستند. مهمترین اسیدهای اشباع نشده ، C_18 میباشند.
روغنهای مایع به علت داشتن پیوندهای Л آسیب پذیرند و لذا با هیدروژندار کردن کاتالیزوری ، پیوندهای دوگانه را از بین میبرند تا نگهداری آنها آسانتر گردد. خیلی از روغنهای جامدی که در آشپزی مورد استفاده قرار میگیرند، از هیدروژندار کردن روغن دانهها و غلات تهیه میشوند. هیدروژندار کردن چربیها ، با اینکه امکان نگهداری این مواد را فراهم میسازد، ولی هضم آنها را در متابولیسم با اشکالی مواجه میسازد.
در سالهای اخیر ، معلوم شده است که این چربیها موجب مسدود شدن رگهای خونی و امراض قلبی میگردند.
روش تهیه استرها و لاکتونها
اولین و قدیمیترین روش سنتز استرها ، واکنش اسیدهای آلی با الکلها در حضور اسید معدنی(معمولا اسید سولفوریک) میباشد. با این روش ، میتوان خیلی از استرها را بطور مستقیم سنتز نمود که این روش ، روش فیشر (Fisher) نامیده میشود. در مواقعی که اسید یا الکل بکار رفته ، ارزان قیمت باشد، میتوان مقدار یکی از واکنشگرها را چند برابر دومی انتخاب کرد و تعادل را به نفع تشکیل محصول بیشتر جابجا کرد و استر بیشتری بدست آورد.
گاهی اوقات برای حصول نتیجه بهتر ، میتوان یکی از محصولات را از محیط خارج کرد و در نتیجه ، واکنش را به سمت تشکیل محصول بیشتر سوق داد.
صابونی شدن
استرها در محلولهای قلیایی آبکی تحت حمله قرار میگیرند و نمک اسید آلی را تولید مینمایند. این واکنش به صابونی شدن موسوم میباشد. برای تائید صحت مکانیسم واکنش از استرهای حاوی اکسیژن با جرم اتمی 18 که یک اتم ایزوتوپ است، استفاده میشود و مشاهده میگردد که اتم اکسیژن سنگین همراه الکل آزاد میشود و نشان میدهد که حمله هسته خواهی به گروه کربونیل استر انجام شده است.
تهیه الکل از استر
استرها با ترکیبات آلی فلزی واکنش میدهند. از این روش برای تهیه الکلهای مربوطه استفاده میشود. از واکنش استرهای فرمیات با ترکیبات آلی فلزی الکل نوع دوم تولید میشود
کربوهیدراتها دستهای از ترکیبات شیمیایی طبیعی بسیار مهم از مواد تشکیل دهنده گیاهان ، گلها ، سبزیجات و درختان هستند. به علاوه کربوهیدراتها به عنوان سیستم منبع ذخیره انرژی عمل میکنند، آنها به آب ، کربن دیاکسید و گرما یا سایر انواع انرژی متابولیزه میشوند
کربوهیدراتها ، منبع مهم غذایی
بدین ترتیب کربوهیدراتها ، منبع مهم غذاییاند. کربوهیدراتها همچنین به عنوان واحدهای سازنده چربیها و نوکلئیک اسیدها عمل میکنند. سلولز ، نشاسته و قند معمولی ، کربوهیدرات میباشند. از آنجا که این ترکیبات چندین گروه عاملی دارند به چند عاملی موسومند. مانند گلوکز و بسیاری از اجزا سازنده ساده ، کربوهیدراتهای پیچیده دارای فرمول عمومی هستند.
نام و ساختار کربوهیدراتها
سادهترین کربوهیدراتها ، قندها یا ساکاریدها هستند. با افزایش طول زنجیر ، تعداد کربنهای با مراکز فضایی افزایش مییابند و بدین ترتیب تعداد زیادی دیاسترومر امکانپذیر میشوند. خوشبختانه برای شیمیدانها عمدتا یکی از چند انانیتومر امکانپذیر ، اهمیت دارد. قندها ترکیبات پلی هیدروکسی کربونیلاند، از این رو میتوانند همی استالهای حلقوی پایداری ایجاد کنند ، بدین ترتیب ساختارهای اضافی و تنوع شیمیایی برای این ترکیبات پدید میآید.
طبقه بندی قندها
کربوهیدراتها نام عمومی قندهای منومری(منوساکاریدها) ، دیمری (دیساکاریدها) ، تریمری(تریساکاریدها) ، الیگومری (الیگوساکاریدها) و پلیمری(پلیساکاریدها) بدست آمده از قند (ساکاروم ، لاتین قند) میباشند. یک منوساکارید یا قند ساده ، یک آلدئید یا کتونی با حداقل دو گروه هیدروکسیل است. بدین ترتیب دو عضو ساده این طبقه از ترکیبات ، 2 و 3 - دی هیدروکسی پروپانال (گلیسرآلدئید) ، 1 و 3 - دی هیدروکسی پروپانون (1 و 3 - دی هیدروکسی استون) میباشند.
قندهای پیچیده از اتصال قندهای ساده همراه با حذف آب بدست میآیند. قندهای آلدئیدی بصورت آلدوزها طبقه بندی میشوند. آنهایی که عامل کتونی دارند، کتوز خوانده میشوند. بر اساس طول زنجیر ، قندها ، تریوز (3 کربنی) ، تتروز(4 کربنی) ، پنتوز (5 کربنی) ، هگزوز (6 کربنی) و غیره نامیده میشوند. از اینرو ، 2 و 3 - دی هیدروکسی پروپانول (گلیسرآلدئید) یک آلدوتریوز است. در حالی که 1 و 3 - دی هیدروکسی پروپانون یک کتوتریوز میباشد.
گلوکز
گلوکز ، قند خون یا قند انگور (گلایکیس ، در فرهنگ یونانی به معنی شیرین) که به دکستروز موسوم است، یک پنتاهیدروکسی هگزانال بوده ، از اینرو در خانواده آلدوزهگزوزها جای دارد. گلوکز بصورت طبیعی در بسیاری از میوهها و گیاهان با غلظتی در گستره %0.08 تا 0.1% در خون انسان وجود دارد.
فروکتوز
ایزومر کتوهگزوزی گلوکز ، فروکتوز است. فروکتوز شیرینترین قند طبیعی است (برخی از قندهای سنتزی شیرینترند). فروکتوز نیز در بسیاری از میوهها (فروکتوز در فرهنگ لاتین به معنی میوه) و در عسل وجود دارد.
ریبوز
قند طبیعی مهم دیگر آلدوپنتزو ریبوز است. این قند واحد ساختاری ریبونوکلئیک اسیدها میباشد. فرمول ساده یا تجربی برای همه قندها میباشد. این فرمول ، هم ارز فرمول هیدرات کربن است. این یکی از دلایلی است که به این دسته از ترکیبات کربوهیدرات گفته میشود.
دیساکاریدها و پلیساکاریدها
دیساکارید از تشکیل دو مونوساکارید از طریق تشکیل یک پل اتری (معمول استال) بدست میآید. هیدرولیز دیساکاریدها ، منوساکاریدها را دوباره بدست میدهد. تشکیل اتر بین یک منو و یک دیساکارید یک تریساکارید ایجاد میکند و تکرار این فرآیند نهایتا به تولید یک پلیمر طبیعی (پلیساکارید) منجر میشود. چنین کربوهیدراتهای پلیمری ، تشکیلدهنده اسکلت اصلی سلولز و نشاسته هستند.
فعالیت نوری قندها
به استثناء 1 و 3 - دیهیدروکسی- پروپانون ، همه قندهایی که تاکنون ذکر شدهاند، حداقل حاوی یک مرکز فضاییاند. سادهترین قند کایرال ، 3 و 2 - دیهیدروکسی پروپانون (گلیسرآلدئید) با یک کربن نامتقارن است. فرم راستبر آن R است، به صورتی که در طرحهای فیشر مولکول نشان داده میشود، انانتیومر چپبر آن ، S میباشد.
گر چه نامگذاری S و R برای نامیدن قندها کاملا رضایت بخش است، اما سیستم نامگذاری قدیمی هنوز بکار گرفته میشود. این سیستم نامگذاری ، قبل از تدوین پیکربندی مطلق قندها متداول بوده ، همه قندها را به 2 و 3 - دیهیدروکسی پروپانال (گلیسرآلدئید) مرتبط میسازد. در این روش بجای استفاده از S و R از پیشوند D برای انانتیومر (+) وLبرای انانتیومر (-) گلیسرآلدئید استفاده میشود.
قندها ، تشکیلدهنده همیاستالهای درون مولکولی
قندها ترکیبات هیدروکسی کربونیلاند و بایستی قادر به تشکیل درون مولکولی همی استال باشند. در واقع گلوکز و سایر هگزوزها و پنتوزها به صورت مخلوط در حال تعادل با ایزومرهای حلقوی همیاستال خود هستند. در این مخلوط در حال تعادل ، ایزومر حلقوی همیاستال برتر است. در اصل هر یک از پنج گروه هیدروکسی میتوانند به گروه کربونیل آلدئید افزوده شوند. اما گرچه حلقههای پنج ضلعی نیز شناخته شده هستند، حلقههای شش ضلعی معمولا محصول برتر میباشند.
گسستگی اکسایشی قندها
واکنشگری که باعث شکستن پیوند C-C میشود، پریدیک اسید (HIO4) است. این ترکیب دیالهای مجاور را اکسایش کرده ، ترکیبات کربونیل ایجاد میشوند. از آنجا که اغلب قندها چندین دیال مجاور دارند، اکسایش با HIO4مخلوط پیچیدهای ایجاد میکند. مقدار کافی از اکسنده ، زنجیر قند را بطور کامل به ترکیبات یک کربنی تبدیل میکند.
از این روش برای شناسایی ساختار قندها استفاده میشود. مثلا از مجاورت گلوکز با 5 اکی والان HIO4، پنج اکی والان فرمی اسید و 1اکی والان فرمالدئید بدست میآیند. در اکسایش فروکتوز ایزومری نیز همان مقدار عامل اکسنده مصرف شده، اما محصولات ، 3 اکی والان اسید ، 2 اکی والان آلدئید و یک اکی والان دیاکسید هستند.
قسمت اول تفسیر آزمایش خون رو براتون تدارک دیدیم که با خوندن اون میتونید با توجه به مقدار موجود هماتوکریت و هموگلوبین خون آنها را تفسیر کنید.
مقدار هموگلوبین و هماتوکریتa
Hgb یا (Hemoglobin)
در برگههای آزمایش مختلف ممکن است به صورتهای مختلف HGB،Hg، یا Hgb نوشته شود. همه این ها مخفف کلمه هموگلوبین، یکی از عناصر اصلی تشکیل دهنده گلبول های قرمز است که با اکسیژن ترکیب می شود و آن را در خون حمل می کند.
این ماده که در آن آهن به کار رفته، جایگاههای مختلفی برای ترکیب با اکسیژن دارد. هموگلوبین در جایی که اکسیژن زیاد وجود دارد با آن ترکیب میشود و در محیط کم اکسیژن، آن را آزاد میکند.
اندازهگیری مقدارکلی هموگلوبین، به نوعی نشانهنده تعداد گلبولهای قرمز است.
مقادیر اصلی مقدار طبیعی : برای آقایان ۱۴ تا ۱۸ گرم در دسیلیتر و برای خانمها مقادیر ۱۲ تا ۱۶ گرم در دسیلیتر.
محدوده خطر: هموگلوبین زیر ۵ و بالای ۲۰ مقادیر خطرزا به حساب میآیند و حتما نیازمند رسیدگی فوری هستند.
چه چیزهایی باعث کاهش هموگلوبین میشود؟
همان دلایلی که باعث کاهش گلبولهای قرمز می شوند، با تخریب هموگلوبین، مقدار آن را هم کم می کنند. کمخونی، خونریزی شدید، سرطان، سوءتغذیه، بیماری لوپوس، بیماریهای کلیوی و بزرگی طحال باعث کاهش هموگلوبین میشوند. مصرف آنتیبیوتیکها، آسپیرین و ایندومتاسین و همچنین داروهای ضدسرطان هم مقدار هموگلوبین خون را کم میکنند
چه چیزهایی باعث افزایش هموگلوبین میشود؟
مشکلات ریوی، سوختگی شدید، نارسایی مزمن ریه و از دست دادن زیاد آب بدن(دهیدراسیون) مقدار این ماده حیاتی را افزایش میدهند. داروهای جنتامایسین و متیلدوپا هم هموگلوبین را افزایش می دهند.
نکته:
- مقدار هموگلوبین در بارداری کاهش مییابد، چون با اینکه خونسازی کمی بیشتر شده است، اما حجم مایع بدن و خون بالا رفته و مقدار کلی هموگلوبین در هر دسیلیتر کاهش مییابد.
- زندگی در ارتفاعات هم به خاطر نیاز بیشتر بدن به اکسیژن و کمبود اکسیژن محیط، باعث تولید بیشتر هموگلوبین میشود.
- در طحال اغلب سلولهای پیر خون تخریب میشوند. بزرگ شدن طحال، یعنی تخریب بیشتر سلولها و به دنبال آن کاهش گلبول های قرمز و هموگلوبین رخ میدهد.
HCt یا (Hematocrit)
هماتوکریت یا HCT هم یکی از مقادیر اندازهگیری گلبول قرمز است. به طور کلی هِم (heme) به معنای آهن است و هر جا در هر کلمهای آمد، حتما آن کلمه ارتباطی با گلبول قرمز دارد.
هماتوکریت درصدی از حجم کل خون است که شامل گلبول های قرمز می باشد و با اندازهگیری قسمت قرمز رسوب خون در لوله آزمایش نسبت به کل ارتفاع خون اندازهگیری میشود.
به خاطر بیماریها و شرایط مختلفی که میتوانند اندازهگیری هموگلوبین و گلبول قرمز را با اشکال مواجه کنند، HCT هم اندازهگیری میشود تا به طور مستقیم نشاندهنده اندازه هموگلوبین و گلبول قرمز باشد. این عدد معمولا با درصد نشان داده میشود.
مقادیر طبیعی : ۴۲ تا ۵۲ درصد برای آقایان و ۳۷ تا ۴۷ درصد برای خانمها. در خانمهای باردار درصد بالاتر از ۳۳ طبیعی است.
محدوده خطر : HCT بالاتر از ۶۰ درصد و پایینتر از ۱۵ درصد.
چه چیزهایی باعث کاهش آن میشود؟
تقریبا همان عواملی که باعث کاهش هموگلوبین و گلبول های قرمز خون می شوند. شرایطی مثل پرکاری تیروئید، سیروز کبدی، نارسایی مغز استخوان و میلوم مولتیپل Multiple myeloma نیز باعث کاهش هماتوکریت میشوند.
چه چیزهایی باعث افزایش آن میشود؟
سوختگی، اسهال شدید، بیماریهای انسدادی ریوی، از دست دادن زیاد آب بدن، تولید بیش از حد گلبول قرمز از عوامل افزایش دهنده ی HCT هستند.
نکته:
- بیماریهایی که باعث به وجود آمدن شکلهای غیرطبیعی گلبول قرمز میشوند (مثل بیماری گلبول قرمز داسیشکل) مقدار HCT را تغییر میدهند.
- وقتی مقدار گلبول سفید خیلی بالا باشد، بر مقدار HCT موثر است.
- در صورت طبیعی بودن اندازههای گلبول قرمز، مقدار هماتوکریت، سه برابر هموگلوبین است.
- هماتوکریت را نباید بلافاصله بعد از خونریزی شدید اندازهگیری کرد.
.: Weblog Themes By Pichak :.
